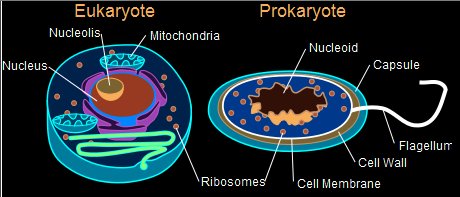
Endosymbiotic
Theory Introduction
 The
hypothesized process by which prokaryotes gave rise to the
first eukaryotic cells is known
as endosymbiosis, and certainly
ranks among the most important evolutionary events. Endosymbiotic
theory, that attempts to explain the origins of eukaryotic
cell organelles such as mitochondria
in animals and
fungi and chloroplasts in plants was
greatly advanced by the seminal work of biologist Lynn Margulis
in the 1960s. Mitochondria are
one of the many different types of organelles in the cells
of all eukaryotes. In general, they are
considered to have originated from proteobacteria (likely
Rickettsiales) through endosymbiosis. Chloroplasts are
one of the many different types of organelles in the plant
cell.
In general, they are
considered to have originated from cyanobacteria through
endosymbiosis. Endosymbiosis has gained ever more acceptance
in the last half century, especially with the relatively
recent advent of sequencing technologies.
There are many variants to the theory, regarding what organism(s)
engulfed what other organism(s), as well as how many times
and when it occurred across geological time. The biology
is messy, and there are many competing theories, so here
we tend to converge them in a unified conceptualization [for
more detailed treatment, visit the "Origins
of the Eukara" pages at Palaeos]. The
hypothesized process by which prokaryotes gave rise to the
first eukaryotic cells is known
as endosymbiosis, and certainly
ranks among the most important evolutionary events. Endosymbiotic
theory, that attempts to explain the origins of eukaryotic
cell organelles such as mitochondria
in animals and
fungi and chloroplasts in plants was
greatly advanced by the seminal work of biologist Lynn Margulis
in the 1960s. Mitochondria are
one of the many different types of organelles in the cells
of all eukaryotes. In general, they are
considered to have originated from proteobacteria (likely
Rickettsiales) through endosymbiosis. Chloroplasts are
one of the many different types of organelles in the plant
cell.
In general, they are
considered to have originated from cyanobacteria through
endosymbiosis. Endosymbiosis has gained ever more acceptance
in the last half century, especially with the relatively
recent advent of sequencing technologies.
There are many variants to the theory, regarding what organism(s)
engulfed what other organism(s), as well as how many times
and when it occurred across geological time. The biology
is messy, and there are many competing theories, so here
we tend to converge them in a unified conceptualization [for
more detailed treatment, visit the "Origins
of the Eukara" pages at Palaeos].
Symbiosis
and Co-evolution
 Symbiosis
is ubiquitous among organisms throughout the tree of life,
from the species level to the kingdom level,
and
even to
the domain level. It is integral to evolution as cooperating
organisms gain survival advantage by a quid pro quo between
them. For example, you (and
for that matter all herbivores omnivores) could not digest
your food without the exquisite symbiosis between your
gut and the
bacteria therein. Symbiosis played a major role in the
co-evolution of flowering plants and the animals that pollinate
them.
The fossil record indicates
that the first flowering plants had primitive flowers.
Through natural selection, adaptive speciation
quickly gave rise to many diverse groups of plants with
specialized, and, at the same time, corresponding speciation
occurred in
certain insect
groups. Many Symbiosis
is ubiquitous among organisms throughout the tree of life,
from the species level to the kingdom level,
and
even to
the domain level. It is integral to evolution as cooperating
organisms gain survival advantage by a quid pro quo between
them. For example, you (and
for that matter all herbivores omnivores) could not digest
your food without the exquisite symbiosis between your
gut and the
bacteria therein. Symbiosis played a major role in the
co-evolution of flowering plants and the animals that pollinate
them.
The fossil record indicates
that the first flowering plants had primitive flowers.
Through natural selection, adaptive speciation
quickly gave rise to many diverse groups of plants with
specialized, and, at the same time, corresponding speciation
occurred in
certain insect
groups. Many  plants are pollinated by insects and vertebrates
(e.g., bats and or birds) that have evolved highly specialized
flowers facilitating pollination by a specific group or
species that are themselves concomitantly adapted through co-evolution.
Such mutualistic associations, where both host and symbiont
evolve to accommodate one another abound in the history
of
life. But,
we digress, so let's return to endosymbiosis. The flower-pollinator
relationship is a common example of symbiosis and resultant
co-evolution. Many flowers have close relationships
with one or a few specific pollinating organisms. Many
flowers, for example, attract only
one specific species of insect, and therefore rely on that insect
for successful reproduction. This close relationship is often
given as an example of coevolution, as the flower and pollinator
are thought to have developed together over a long period of
time to match each other's needs. This close relationship compounds
the negative effects of extinction. The extinction of either
member in such a relationship would
mean almost certain extinction of the other member as well. Some
endangered plant species are so because of shrinking pollinator
populations. plants are pollinated by insects and vertebrates
(e.g., bats and or birds) that have evolved highly specialized
flowers facilitating pollination by a specific group or
species that are themselves concomitantly adapted through co-evolution.
Such mutualistic associations, where both host and symbiont
evolve to accommodate one another abound in the history
of
life. But,
we digress, so let's return to endosymbiosis. The flower-pollinator
relationship is a common example of symbiosis and resultant
co-evolution. Many flowers have close relationships
with one or a few specific pollinating organisms. Many
flowers, for example, attract only
one specific species of insect, and therefore rely on that insect
for successful reproduction. This close relationship is often
given as an example of coevolution, as the flower and pollinator
are thought to have developed together over a long period of
time to match each other's needs. This close relationship compounds
the negative effects of extinction. The extinction of either
member in such a relationship would
mean almost certain extinction of the other member as well. Some
endangered plant species are so because of shrinking pollinator
populations.
Endosymbiotic
Theory and Eukaryotic Origins
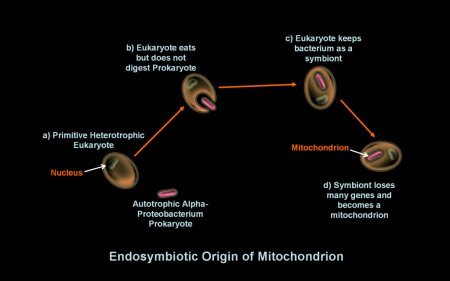 Such
symbiotic relationships in which two species are dependent
upon one another to varying extents also served as crucial
elements of the evolution of eukaryotic cells. The
theory holds that the eukaryote mitochodrion evolved from a
small, autotrophic
bacterium that was engulfed by a larger
primitive, heterotrophic, eukaryotic cell. This eukaryotic cell
arose when an Such
symbiotic relationships in which two species are dependent
upon one another to varying extents also served as crucial
elements of the evolution of eukaryotic cells. The
theory holds that the eukaryote mitochodrion evolved from a
small, autotrophic
bacterium that was engulfed by a larger
primitive, heterotrophic, eukaryotic cell. This eukaryotic cell
arose when an
anaerobic prokaryote (unable to
use oxygen for energy) lost its cell wall. The more flexible
membrane underneath then began to grow and fold in on itself
which, in turn, led to
formation of a nucleus and other internal membranes. Endosymbiosis
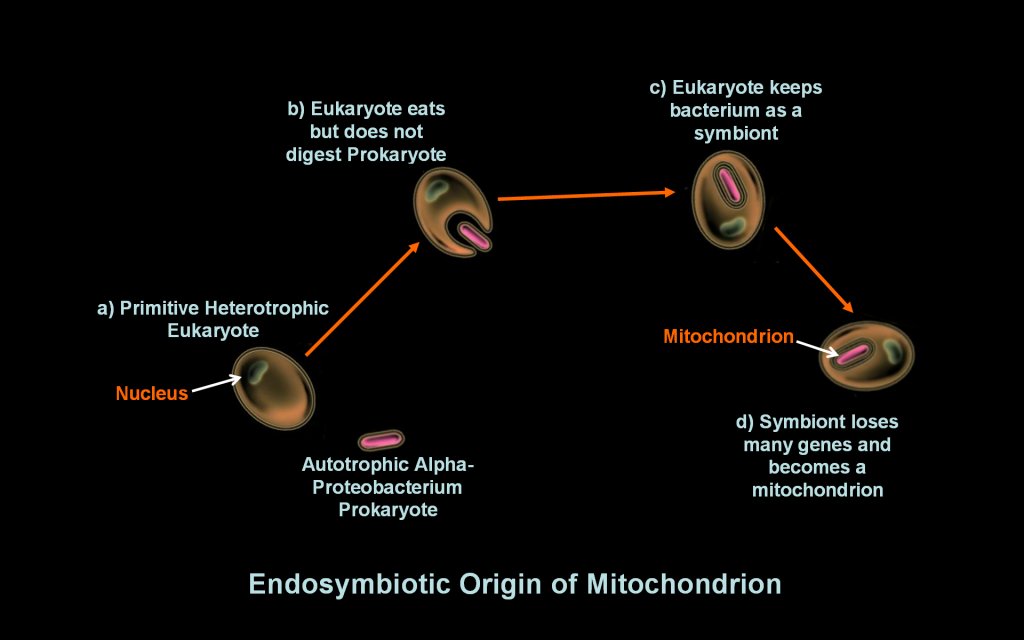
occurred according to the figure to the right: a) The primitive
eukaryotic cell was also eventually able to eat
prokaryotes, a marked improvement to absorbing small
 molecules
from its environment; b) The process of endosymbiosis commenced
when the eukaryote engulphed but did not digest a autotrophic
bacterium. Evidence suggests this engulfed bacterium was an alphaproteobacteria,
an autotroph that uses photosynthesis
to acquire energy. c) The eukaryote then began a mutually beneficial
(symbiotic) relationship with it whereby the eukaryote provided
protection and nutrients to the prokaryote, and in return, the
prokaryotic endosymbiont provided additional energy to its eukaryotic
host
through its
respiratory cellular machinary. d) The relationship became permanent
over time completing primary endosymbiosis as the endosymbiont
lost some genes it used for independent life and transferred
others to
the eukaryote's
nucleus. The symbiont thus became dependent
on the host cell for organic molecules
and
inorganic compounds. The genes of the repiratory machinary
became a
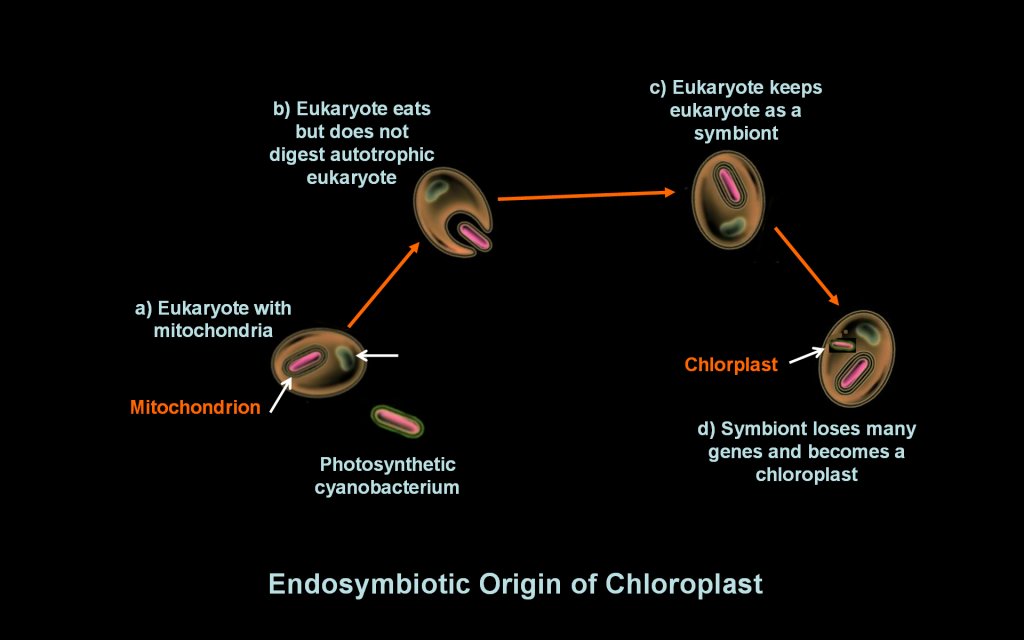
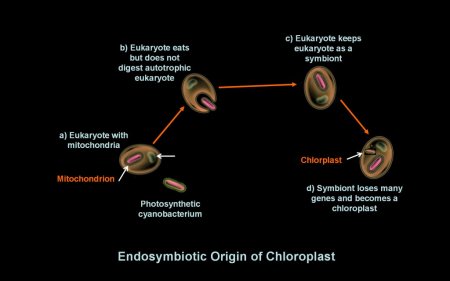
mitochondrion. Endosymbiotic theory hypothesizes the origin of
chloroplasts similarly, where chloroplasts a eukaryote with mitochondria
engulfs a photosynthetic cyanobacteruim in a symbiotic relationship
ending
in the chloroplast organelle. molecules
from its environment; b) The process of endosymbiosis commenced
when the eukaryote engulphed but did not digest a autotrophic
bacterium. Evidence suggests this engulfed bacterium was an alphaproteobacteria,
an autotroph that uses photosynthesis
to acquire energy. c) The eukaryote then began a mutually beneficial
(symbiotic) relationship with it whereby the eukaryote provided
protection and nutrients to the prokaryote, and in return, the
prokaryotic endosymbiont provided additional energy to its eukaryotic
host
through its
respiratory cellular machinary. d) The relationship became permanent
over time completing primary endosymbiosis as the endosymbiont
lost some genes it used for independent life and transferred
others to
the eukaryote's
nucleus. The symbiont thus became dependent
on the host cell for organic molecules
and
inorganic compounds. The genes of the repiratory machinary
became a
mitochondrion. Endosymbiotic theory hypothesizes the origin of
chloroplasts similarly, where chloroplasts a eukaryote with mitochondria
engulfs a photosynthetic cyanobacteruim in a symbiotic relationship
ending
in the chloroplast organelle.
When
these endosymbiotic events occured
is subject to much debate, but it must have been early
in life's history, perhaps as early as the
Archean Eon more than 2500 million years ago. The
heterotrophic prokaryote
used cellular respiration to intake oxygen and convert
organic
molecules
to energy. The prokaryotic cells that were too small
to be digested continued to live inside the host eukaryote,
eventually
becoming
dependent on the host cell for organic molecules and
inorganic compounds. Importantly, the host cell could have
acquired,
by the addition of the aerobic function, an increased
output of
ATP for cellular activities, leading to its improved
selective advantage. Was the "engulfer" a eubacteria
or an archaean - yes - it depends on which of competing
theories
you choose?
Other
theories hold that the prokaryotes that gave rise to
early eukaryotes
were probably from the Domain Archaea,
both because of several key characteristics and because
DNA sequence comparison suggest that
archaeans are more closely related to the eukaryotes
than are eubacteria. This is the so-called serial endosymbiosis
theory
of a monophyletic origin of the mitochondrion from
a eubacterial ancestor. That fact that mitochondria
have their own DNA, RNA,
and ribosomes, supports the endosymbiosis theory, as
does the existence of the amoeba, a eukaryotic organism
that lacks mitochondria
and therefore requires a symbiotic relationship with
an aerobic bacterium.
Endosymbiosis
Leads to Mitochondria
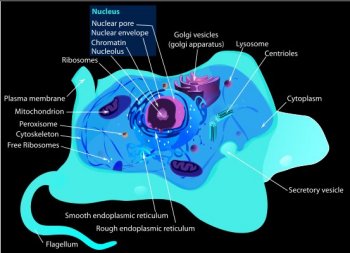 Digging
deeper, the symbiosis is analogous to that between plants and
their "birds and bees"
symbionts. The aerobic bacterium thrived
within the cell cytoplasm that provided abundant molecular
food for its heterotrophic existence. The bacterium digested
these
molecules that manufactured enormous energy in the form of
adenosine triphosphate (ATP), and so much so that extra ATP
was available to the host cell's cytoplasm. This
enormously benefited the anaerobic cell that then gained the
ability to
aerobically
digest
food. Eventually,
the aerobic bacterium could no longer live independently from
the cell, evolving into the mitochondrion organelle. Such aerobically
obtained energy vastly exceeded that of anaerobic respiration,
setting the
stage for vastly accelerated evolution of eukaryotes. Digging
deeper, the symbiosis is analogous to that between plants and
their "birds and bees"
symbionts. The aerobic bacterium thrived
within the cell cytoplasm that provided abundant molecular
food for its heterotrophic existence. The bacterium digested
these
molecules that manufactured enormous energy in the form of
adenosine triphosphate (ATP), and so much so that extra ATP
was available to the host cell's cytoplasm. This
enormously benefited the anaerobic cell that then gained the
ability to
aerobically
digest
food. Eventually,
the aerobic bacterium could no longer live independently from
the cell, evolving into the mitochondrion organelle. Such aerobically
obtained energy vastly exceeded that of anaerobic respiration,
setting the
stage for vastly accelerated evolution of eukaryotes.
Endosymbiosis
Leads to Chloroplasts
 Endosymbiotic
theory
posits a later parallel origin of the chloroplasts; a cell
ate
a photosynthetic
cyanobacterium and failed to digest it. The cyanobacterium
thrived in the cell and eventually evolved into the first
chloroplast. Other eukaryotic organelles may have also evolved
through
endosymbiosis; it has been proposed that cilia, flagella,
centrioles, and
microtubules
may have originated from a symbiosis between a Spirochaete
bacterium and an early eukaryotic cell, but this is not
yet broadly accepted
among biologists. Endosymbiotic
theory
posits a later parallel origin of the chloroplasts; a cell
ate
a photosynthetic
cyanobacterium and failed to digest it. The cyanobacterium
thrived in the cell and eventually evolved into the first
chloroplast. Other eukaryotic organelles may have also evolved
through
endosymbiosis; it has been proposed that cilia, flagella,
centrioles, and
microtubules
may have originated from a symbiosis between a Spirochaete
bacterium and an early eukaryotic cell, but this is not
yet broadly accepted
among biologists.
Secondary
Endosymbiosis
Primary
endosymbiosis involves the engulfment of a bacterium by another
free living organism. Secondary endosymbiosis occurs
when the product of primary endosymbiosis is itself engulfed
and retained by another free living eukaryote. Secondary endosymbiosis
has occurred several times and has given rise to extremely diverse
groups of algae and other eukaryotes. Some organisms can take
opportunistic advantage of a similar process, where they engulf
an alga and use the products of its photosynthesis, but once
the prey item dies (or is lost) the host returns to a free living
state. Obligate secondary endosymbionts become dependent on their
organelles and are unable to survive in their absence. The process
of secondary endosymbiosis left its evolutionary signature within
the unique topography of plastid membranes.
Secondary plastids are surrounded by three (in euglenophytes
and some dinoflagellates) or four membranes (in haptophytes,
heterokonts, cryptophytes, and chlorarachniophytes). The two
additional membranes are thought to correspond to the plasma
membrane of the engulfed alga and the phagosomal membrane of
the host cell. The endosymbiotic acquisition of a eukaryote cell
is represented in the cryptophytes; where the remnant nucleus
of the red algal symbiont (the nucleomorph) is present between
the two inner and two outer plastid membranes. Despite
the diversity of organisms containing plastids, the morphology,
biochemistry, genomic organisation,
and molecular
phylogeny of plastid RNAs and proteins suggest a single origin
of all extant plastids – although this theory is still
debated. Some
species including Pediculus humanus have multiple chromosomes in
the mitochondrion. This and the pylogenetics of the genes
encoded within the mitochondrion suggests that the ancestors
of mitochondria may have been acquired on several occasions rather
than just once.
Mitochondria
and Chloroplasts
Cell Powerhouses
We
could fairly posit that the evolutionary
origin of the eukaryotic cell was "the
first time that what went around came around", a quid pro
quo with among primitive organisms in deep time. This would make
the all eukaryotes chimaeras at a cellular level. The Eukaryotic
cell could also be likened to the V8 engine in producing power,
as compared to a donkey powering prokaryotic cells. This would
have enormous implication for subsequent evolution as earth's
oceans atmosphere were oxygenated by photosynthetic bacteria
creating extensive stromatolitic
reefs. Eukaryotes became multicellular in the precambrian at
the same time earth's oxygen levels were rising. More oxygen
together with mitochondria to burn it in was one driving function
for the forthcoming Cambrian
Explosion when the ancestors
of modern
eukaryotes' appeared.
Mitochondria, the result of endosymbiosis in eukaryotic evolution
are the energy-generating V8 engines of eukaryotic cells, where
oxidative phosphorylation and electron transport metabolism takes
place. Plastids, including chloroplasts, are the corresponding
photosynthetic organelles of plant and algae cells.
Mitochondrial
DNA and Function
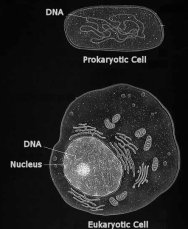 The
mitochondrion is different from most other organelles because
it has its own circular DNA (similar to the DNA of prokaryotes)
and reproduces independently of the cell in which it is found,
one of the major pieces of evidence supporting endosymbiosis.
Although most DNA is packaged in chromosomes within the nucleus,
mitochondria
also
have a
small amount of
their own DNA. This
genetic material is known as mitochondrial DNA or mtDNA. Mitochondria
are
structures
within cells that convert the energy
from food into a form that cells can use.
Each cell contains hundreds to thousands of mitochondria, which
are located in the fluid that surrounds the nucleus (the cytoplasm).
Mitochondria produce energy through a process called oxidative
phosphorylation.
This process uses oxygen and simple sugars to
create adenosine triphosphate (ATP), the cell’s main energy
source. A set of enzyme complexes, designated as complexes I-V,
carry out oxidative phosphorylation within mitochondria. In addition
to energy production, mitochondria play a role in several other
cellular activities. For example, mitochondria
help regulate the self-destruction of cells (apoptosis). They
are also necessary for the production of substances such as cholesterol
and heme (a component of hemoglobin, the molecule that carries
oxygen in the blood). Human mitochondrial DNA contains 37 genes,
all of which are essential for normal mitochondrial function.
Thirteen
of these genes provide
instructions for making enzymes involved in oxidative phosphorylation.
The remaining genes provide instructions for making molecules
called transfer RNAs (tRNAs) and ribosomal RNAs (rRNAs), which
are chemical cousins of DNA. These types of RNA help assemble
protein building blocks (amino acids) into functioning proteins. The
mitochondrion is different from most other organelles because
it has its own circular DNA (similar to the DNA of prokaryotes)
and reproduces independently of the cell in which it is found,
one of the major pieces of evidence supporting endosymbiosis.
Although most DNA is packaged in chromosomes within the nucleus,
mitochondria
also
have a
small amount of
their own DNA. This
genetic material is known as mitochondrial DNA or mtDNA. Mitochondria
are
structures
within cells that convert the energy
from food into a form that cells can use.
Each cell contains hundreds to thousands of mitochondria, which
are located in the fluid that surrounds the nucleus (the cytoplasm).
Mitochondria produce energy through a process called oxidative
phosphorylation.
This process uses oxygen and simple sugars to
create adenosine triphosphate (ATP), the cell’s main energy
source. A set of enzyme complexes, designated as complexes I-V,
carry out oxidative phosphorylation within mitochondria. In addition
to energy production, mitochondria play a role in several other
cellular activities. For example, mitochondria
help regulate the self-destruction of cells (apoptosis). They
are also necessary for the production of substances such as cholesterol
and heme (a component of hemoglobin, the molecule that carries
oxygen in the blood). Human mitochondrial DNA contains 37 genes,
all of which are essential for normal mitochondrial function.
Thirteen
of these genes provide
instructions for making enzymes involved in oxidative phosphorylation.
The remaining genes provide instructions for making molecules
called transfer RNAs (tRNAs) and ribosomal RNAs (rRNAs), which
are chemical cousins of DNA. These types of RNA help assemble
protein building blocks (amino acids) into functioning proteins.
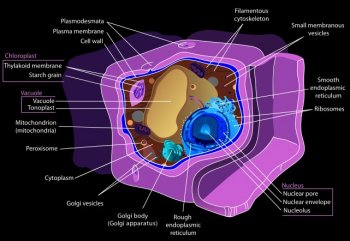
Chloroplast
DNA and Function
Like
mitochondria, chloroplasts also contain their own DNA and are
able to grow and reproduce independently within the cells of
plants and other protista that conduct the complex
chemistry in photosynthesis. In green plants, chloroplasts are
surrounded by two lipid-bilayer membranes thought to correspond
to the outer and inner membranes of the ancestral cyanobacterium
from which chloroplasts descended. Chloroplasts have their
own genome, which is considerably reduced compared to that
of free-living
cyanobacteria,
but the parts that are still present show clear similarities
with the cyanobacterial genome. Plastids typically contain
some 60 to 100 genes, compared cyanobacteria that have some
1500 genes. Many of the apparently missing genes are encoded
in the nuclear
genome
of the host. Chloroplasts capture light energy to conserve free
energy in the form of ATP and reduce NADP to NADPH through
a complex set of processes called photosynthesis.
Evidence
for Endosymbiotic Theory
- Mitochondria have very similar characteristics
to purple-aerobic bacteria. They both use oxygen in the
production of ATP, and
they both do this by using the Kreb’s Cycle and oxidative
phosphorylation. Similarly, chloroplasts are very similar to
photosynthetic bacteria in that they both have similar
chlorophyll
that harnesses light energy that is converted into chemical
energy. Although there are many similarities between mitochondria
and purple
aerobic bacteria and chloroplasts and photosynthetic
bacteria, they appear to be slight and explainable by subsequent
evolution.
- Mitochondria and chloroplasts are similar in size to bacteria,
1 to 10 microns.
- Mitochondria
and chloroplasts DNA, RNA,
ribosomes, chlorophyll (for chloroplasts), and protein synthesis
is similar to that for bacteria. This provided the first
substantive evidence for the endosymbiotic hypothesis. It
was also determined
that mitochondria and chloroplasts
divide independently of the cell they live in. Mitochondria
having their own DNA and dividing independently of the cell
is what
ultimately results in only mitochondrial
DNA being inherited by one’s mother since only an egg
cell has DNA while a sperm cell does not.
- Both
mitochondria and chloroplasts have double phospholipid bilayers.
This appears to have arisen by mitochondria
and chloroplasts entering eukaryotic cells via endocytosis. Both
purple, aerobic bacteria (similar to mitochondria) and photosynthetic
bacteria (similar to chloroplasts) only have one phospholipid
bilayer, but when they enter another cell via endocytosis, they
are bound by a vesicle which forms the second layer of their
double phospholipid bilayer.
|